ABSTRACT: Although crucial to our understanding of skeletal evolution in marine invertebrates, the cost of calcification has remained elusive for a simple reason: calcium carbonate is an inorganic material. Its cost thus derives solely from the metabolic expenses of accumulating, transporting, and precipitating CaCO3, and cannot normally be separated from other metabolic costs. Traditionally, calcification cost has been ignored and total shell cost assumed to derive solely from skeletal organic matrix. The cost estimated here was permitted by the substantial natural variation in shell thickness in two rocky-shore gastropods (Nucella lamellosa and N. lapillus). In both the field and laboratory, data from three separate experiments revealed that groups of snails producing extra shell material under a particular set of experimental conditions also consumed extra food. The cost of calcification was estimated by computing the extra energy assimilated per unit extra shell produced at a common rate of tissue growth, and then subtracting the cost of the organic matrix. At 1-2 J/mg CaCO3, the calcification cost reported here is roughly 5% of that for the predominantly proteinaceous organic fraction of molluscan shells on a per-gram basis. This may explain why calcareous microstructures high in organic content have become less common evolutionarily.
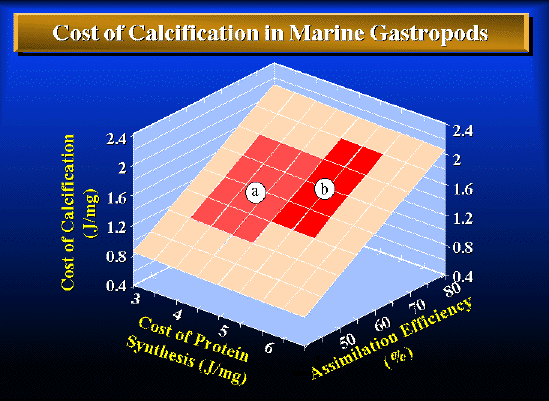
Figure 1. An illustration of how sensitive the estimated cost of calcification is to assumptions I had to make about a) assimilation efficiencies (energy assimilated/energy ingested) and b) the metabolic cost of protein synthesis (J/mg protein), neither of which are known precisely. The relationship describing the surface in the figure is: C= (xI - yP - O)/ (1 - P) where C= cost of precipitating one mg CaCO3 (J/mg), x= assimilation efficiency, y= cost of synthesizing 1 mg of organic matrix (J/mg), I= extra energy ingested/mg shell produced (3.196 J/mg from equation 9, Table 2), P= mg organic matrix/mg shell [0.0166 from Palmer (29)], and O= energy content of organic matrix/mg shell [0.393 J, assuming 1.66% organic matrix (29), and 23.66 J/mg protein (41) since the organic matrix of molluscan shells is predominantly protein with a small fraction of amino sugars (42, 43)]. Region (a) bounds the cost of calcification based upon some less restrictive assumptions: assimilation efficiencies for carnivorous marine gastropods range from 50 - 70% (28) and estimates of the metabolic cost of protein synthesis range from 3.6 - 5.0 J/mg (44-46). Region (b) bounds the cost of calcification based upon more restrictive assumptions for Nucella. The average gross growth efficiency (energy in somatic growth/energy ingested) of 38.8 ± 1.79% [computed across all experiments from data in Table 1 using 24.47 J/mg ash free dry weight for Nucella tissue (38)] suggests that the estimated assimilation efficiency of 65.8 ± 12.3% for Nucella feeding on mussels (27) is reasonable, though perhaps a bit low. Waterlow (46) argues cogently that the estimated metabolic cost of protein synthesis of 3.6 J/mg, based on the cost of peptide bonds, is almost certainly too low. Allowing for costs of amino-acid and protein transport, signal peptides, and maintenance of ribosomal machinery, a 50% increase in the cost of synthesis to 5.4 J/mg may be more reasonable.
A.R. Palmer Publications List, Asymmetry Page, Home Page
Original material on this page copyright © A. Richard Palmer. All rights reserved.
(revised Nov. 4, 2002)