
Heptageniidae of the World: Part II. Key to the Genera
Introduction
In Part I of the Heptageniidae of the World (Ephemeroptera), Wang and McCafferty (2004) analyzed the generic relationships and presented a phylogenetic classification of the family. Three subfamilies, 12 tribes, and 29 genera were recognized. The present paper provides worldwide keys and diagnoses for larvae and male adults of the genera recognized in Part I as well as subsequently described genera and revisions of generic concepts, and a key for female adults of North American genera. The forthcoming Part III will be a catalog of the Heptageniidae with coverage of all valid names, nomenclatural history, and pertinent taxonomic literature (Webb and McCafferty in prep.).
Generic Changes
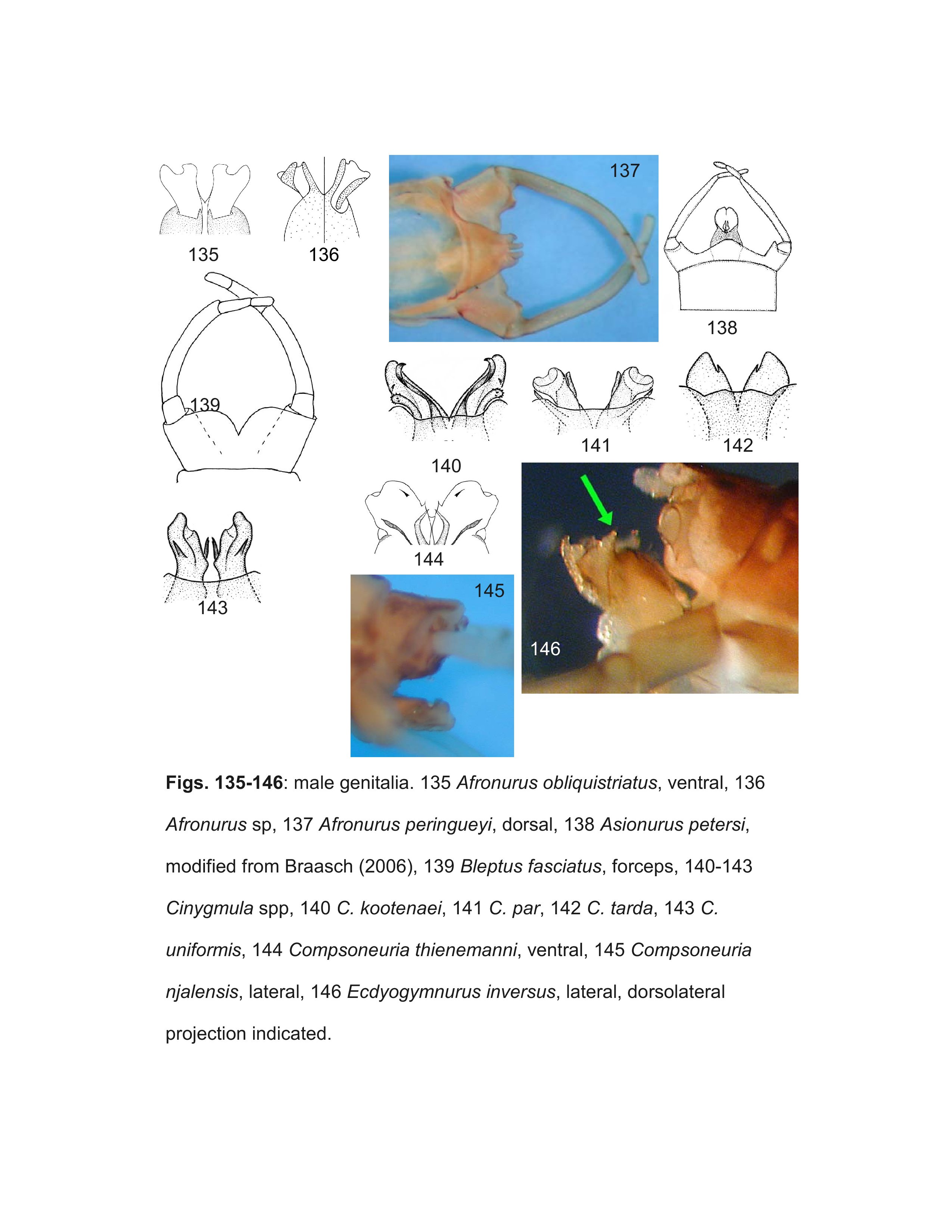
Kluge (2004) established Ecdyogymnurus Kluge as a subgenus of Ecdyonurus Eaton, designating Ecdyonurus inversus Kluge as the type species. Independently, McCafferty (2004) described Siberionurus McCafferty as a new genus, also designating E. inversus as the type species. Because Ecdyogymnurus was published shortly before Siberionurus, it is the senior name of the genus by priority (Ecdyogymnurus Kluge = Siberionurus McCafferty, N. OBJ. SYN.) and we treat it at the generic level herein.
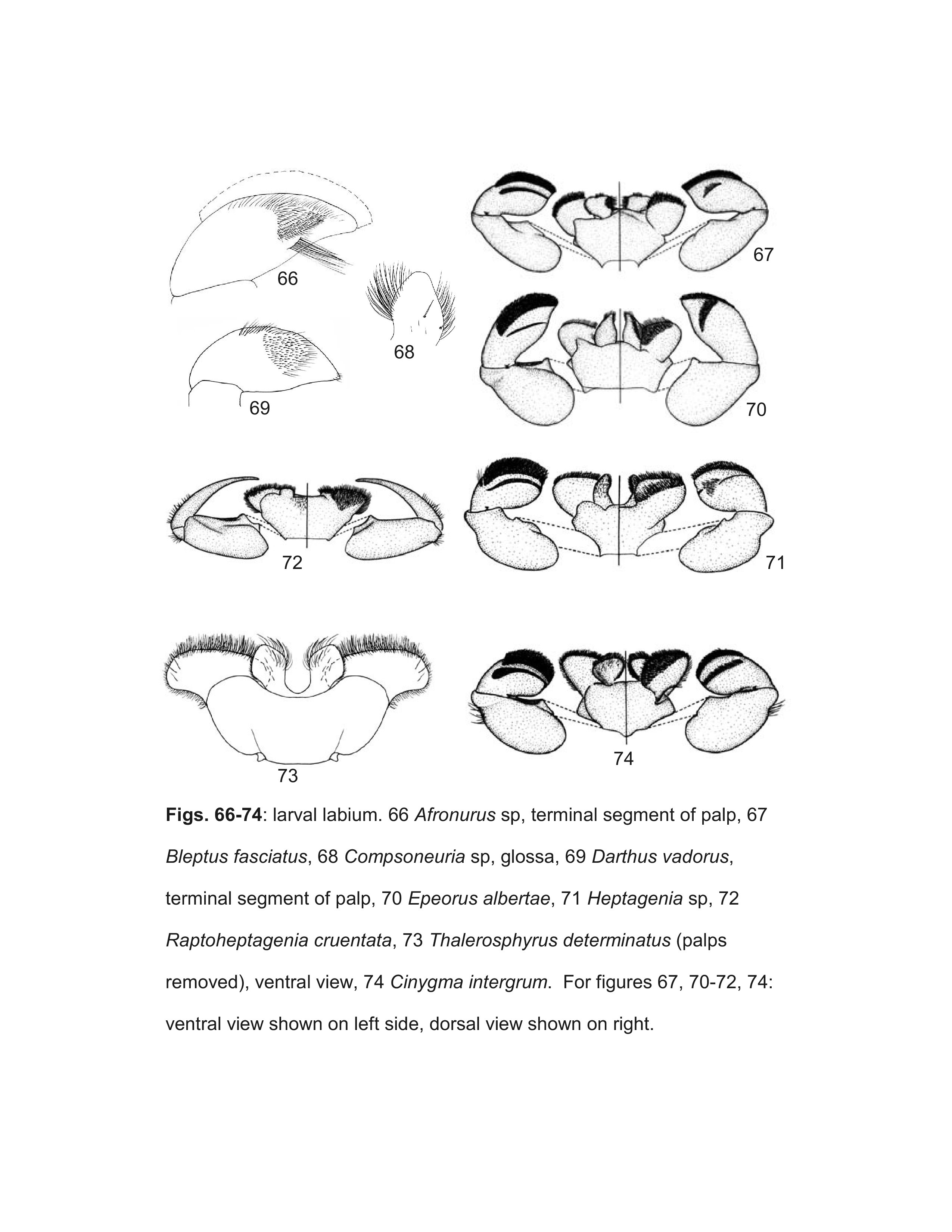
Two genera were described subsequent to Wang and McCafferty's (2004) revision: Parafronurus Zhou & Braasch and Darthus Webb & McCafferty. These two genera were found to belong to a clade with Afronurus Lestage (Webb and McCafferty 2007) and, in accordance with sequencing conventions, the tribe Afronurini Webb & McCafferty was established in order to retain the generic status of Atopopus Eaton, Thalerosphyrus Eaton and Asionurus Braasch & Soldán.
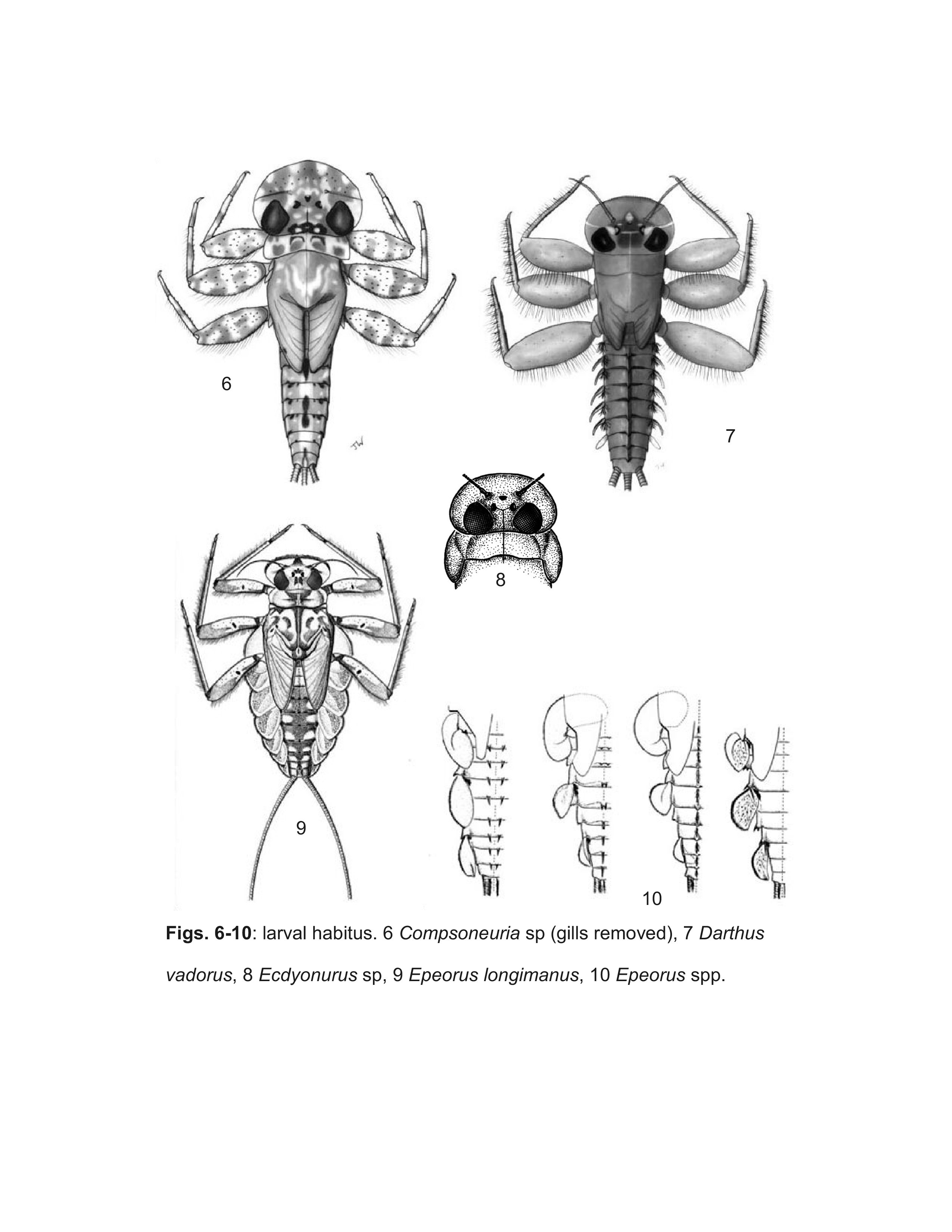
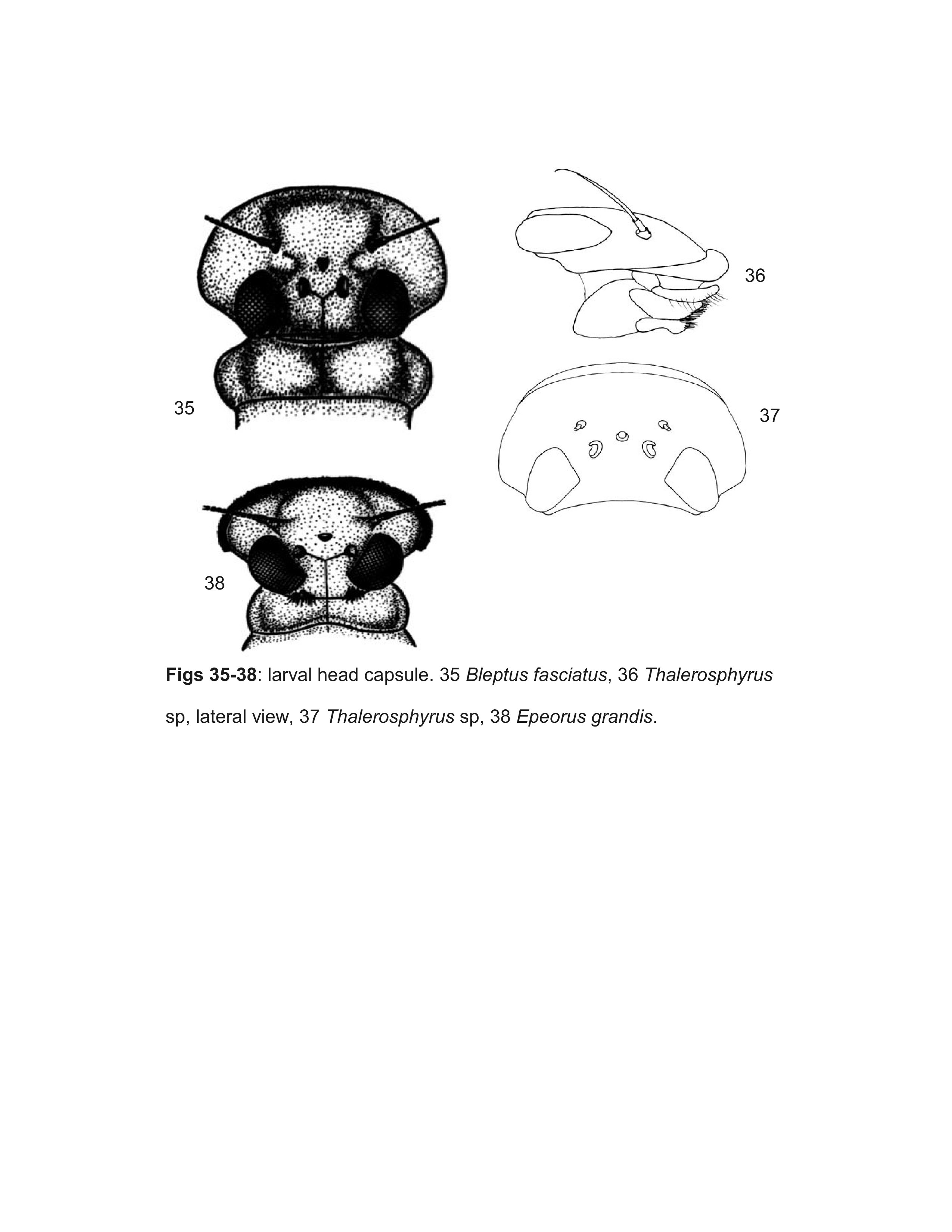
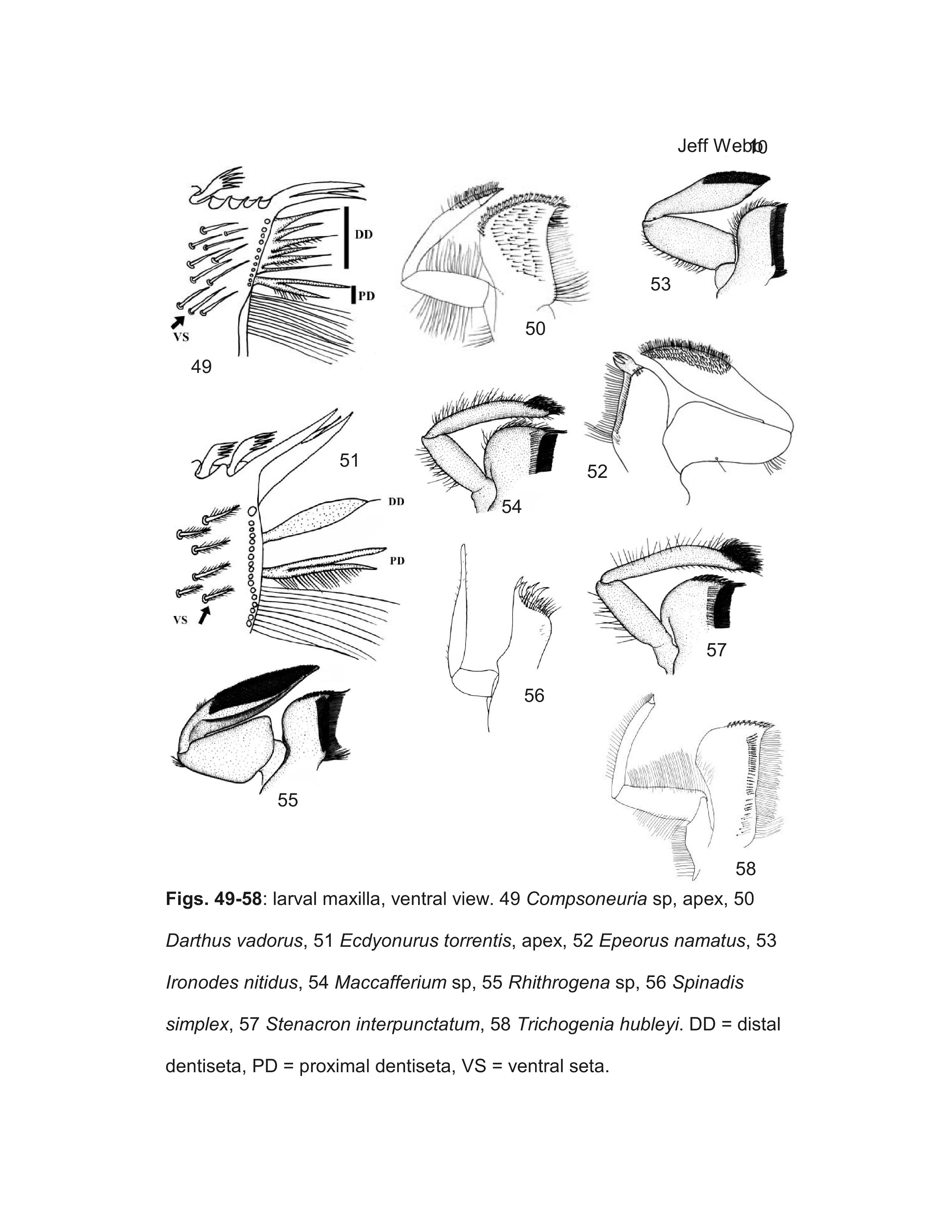
Webb et al. (2006) showed that larvae Ulmer (1939) described as Compsoneuria Eaton are actually a species of Trichogenia Braasch & Soldán. Based on characteristics of adult males, Webb et al. (2006) furthermore showed Compsoneuria to be a member of the Ecdyonurinae and transferred the genus and the tribe Compsoneuriini to that subfamily, at the same time establishing the tribe Trichogeniini Webb & McCafferty in the Heptageniinae for the genus Trichogenia. Webb et al. (2006) additionally recognized the synonymy of Compsoneuriella Ulmer and Notonurus Crass with Compsoneuria, and restricted the concept of Thalerosphyrus.
Wang and McCafferty (2004) placed Epeiron Demoulin as a synonym of Cinygmula because they did not detect any significant difference in the male genitalia, tarsal segment ratios, or thoracic structure between the two genera. All species of Cinygmula possess median and usually ventral titillators, but no species described in Epeiron, including the type species E. amseli Demoulin, have either median or ventral titillators (Demoulin 1964, Kluge 2004). The lack of titillators indicates that Epeiron is equivalent to Rhithrogena, which always lack median titillators (ventral titillators may be present or absent). Although no species initially described in Epeiron are known as larvae, Kluge (2004) showed that several species described in Rhithrogena form a monophyletic group with Epeiron and many of these species are known in the larval stage, such as R. binerve Kluge, R. eugeniae Kluge, R. paulinae Sartori & Sowa, and R. znojkoi Tshernova. These larvae all possess gill lamellae that form a friction disc, characteristic of Rhithrogena. Epeiron was synonymized with Rhithrogena by Kluge (1988) and Sartori and Sowa (1992), and we follow that interpretation herein.
Kluge (2004) recognized five subgenera within Rhithrogena: Himalogena Kluge, Tumungula Zhou & Peters, Sibirigena Kluge, Epeiron, and Rhithrogena. Tumungula, Sibirigena and Epeiron each possess autapomorphies, but Himalogena and Rhithrogena are defined by plesiomorphic characteristics (Kluge 2004). Recognition of thse groupings results in a paraphyletic classification and therefore, we do not recognize any of the subgenera of Rhithrogena (=Himalogena Kluge, N.SYN.; Sibirigena Kluge, N.SYN.; Tumungula Zhou & Peters, N.SYN.). For similar reasons, we do not recognize subgenera of Epeorus (=Alpiron Braasch, N.SYN.; Albertiron Kluge, N.SYN.; Belovius Tshernova, N.SYN.; Caucasiron Kluge, N.SYN.; Iron Eaton; Ironopsis Traver; Proepeorus Kluge, N.SYN.) or Compsoneuria (=Siamoneuria Braasch, N.SYN.).
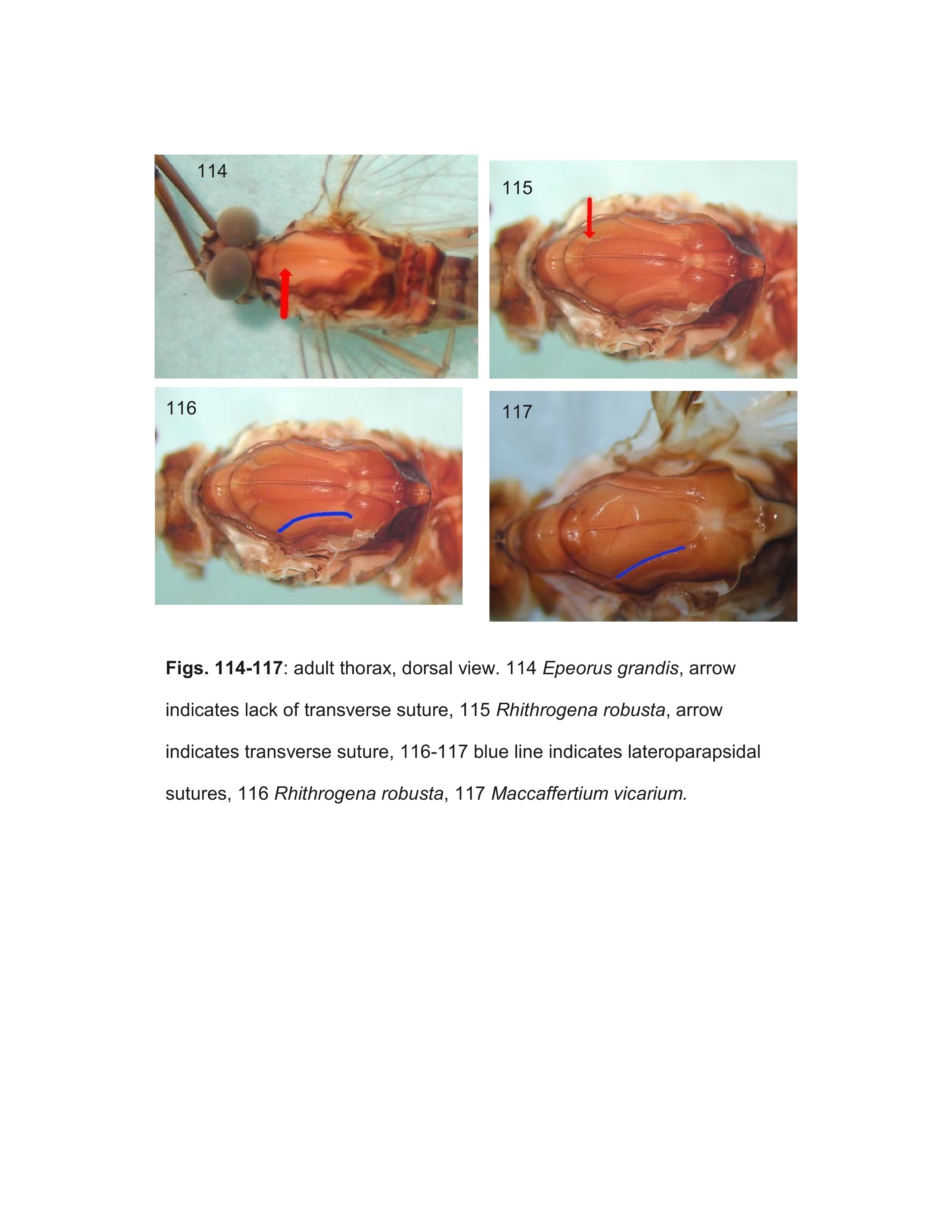
Kluge (2004) established several subgenera within Ecdyonurus, many of which are treated as genera Wang and McCafferty (2004). One of these subgenera, Thamnodontus Kluge, is not defined by any apomorphies, but the three included species cannot be confidently placed in any other genus. Thamnodontus appears to be most similar to Notacanthurus as adults have slightly to greatly expanded head capsules (Kluge 1983). Larvae, however, lack the characteristic median ridge of spines on the abdomen. Even though the monophyly of Thamnodontus is not supported, we are tentatively recognizing it as a valid genus until further evidence becomes available.
Notes on the Keys
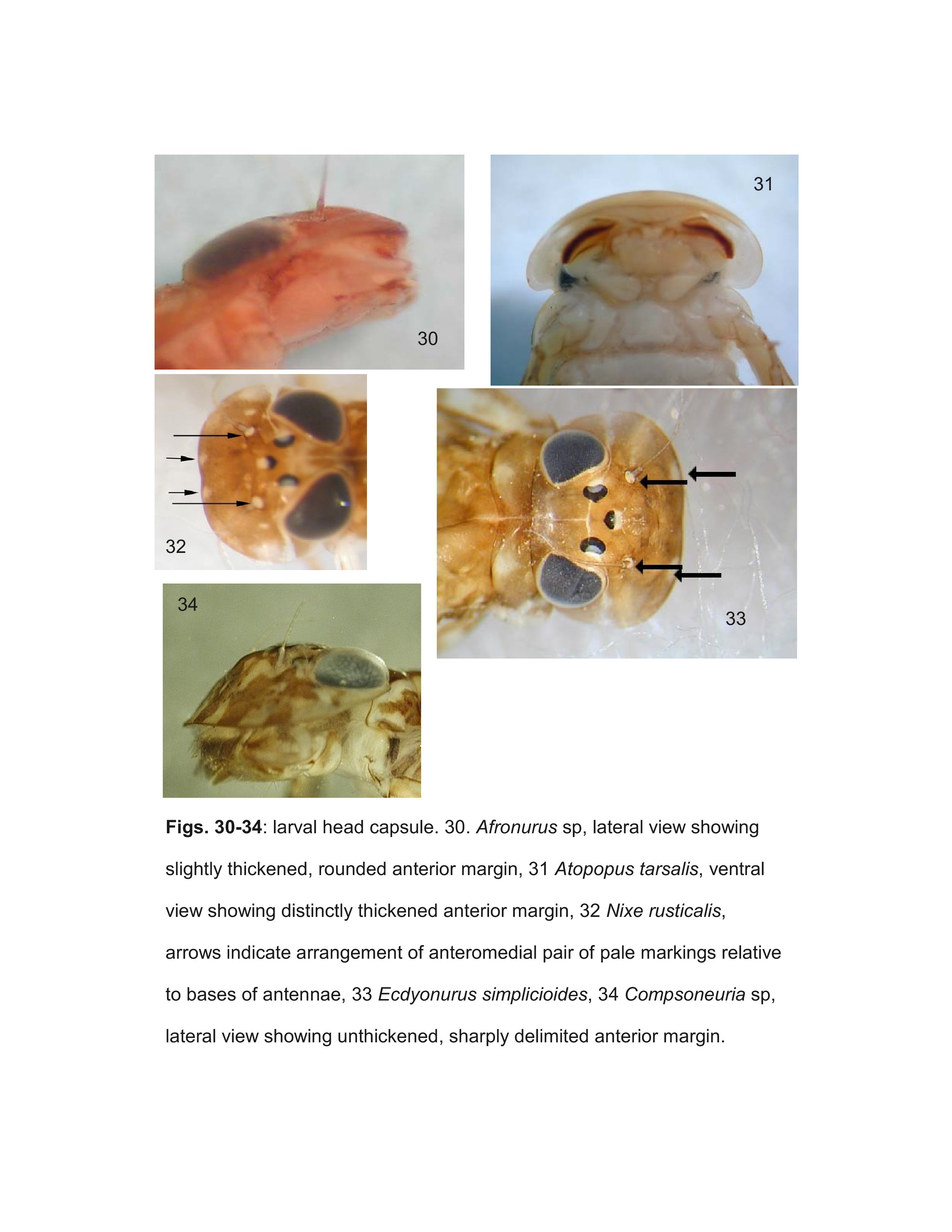
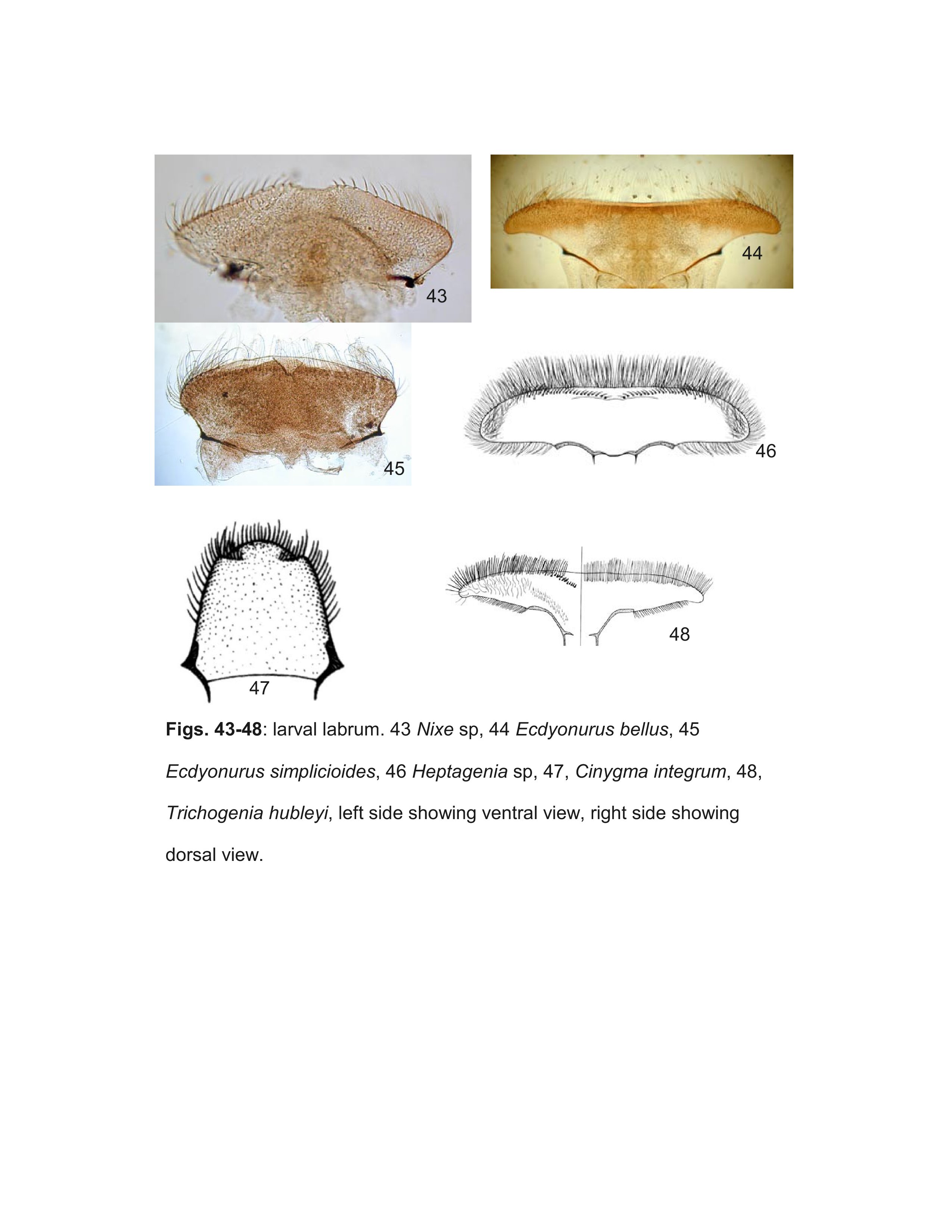
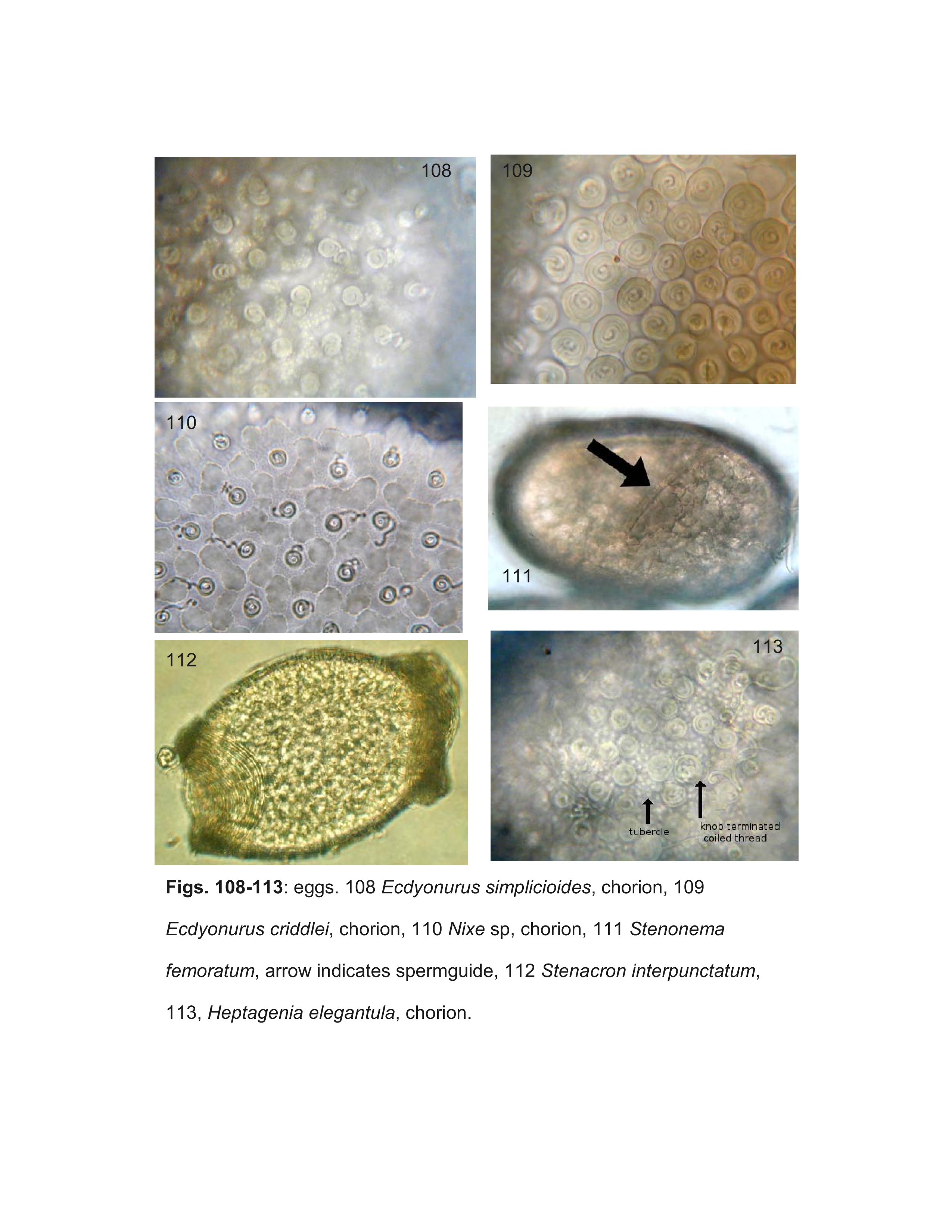
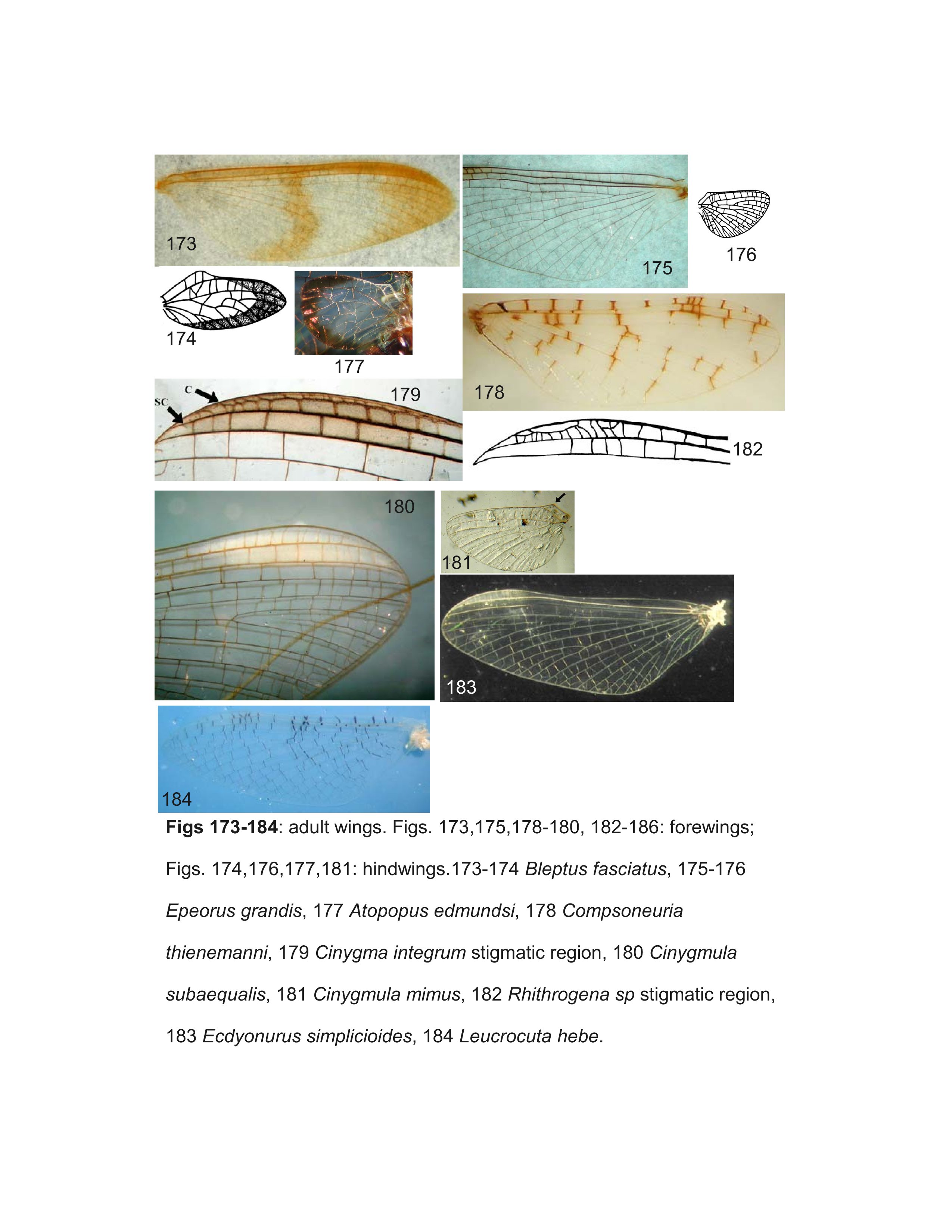
Most specimens of Heptageniinae and Rhithrogeninae should be readily identified using this key. Differentiation of some Ecdyonurinae genera, however, is more provisional, especially in the poorly known Oriental Region. For larvae of Ecdyonurinae, the relative development of the anterior margin of the head capsule is frequently useful, but may be difficult to interpret. The more primitive genera generally have the dorsal and ventral surfaces of the anterior margin distinctly differentiated (Fig. 34). Afronurus, Darthus, and Parafronurus differ in having a rounded anterior margin where the dorsal and ventral surfaces do not meet in a distinct edge (Fig. 30). Thalerosphyrus and Atopopus have the anterior margin distinctly thickened ventrally, and sometimes dorsally as well (Figs. 31, 36). Larvae of Nixe and the Ecdyonurus simplicioides group cannot reliably be distinguished except by examination of eggs from mature females. In North America, Ecdyonurus simplicioides (McDunnough, 1924a) is known to be sympatric with several Nixe spp. in central Canada, but can be distinguished by the colour pattern of the head capsule, as indicated in the key. Nixe kennedyi (McDunnough, 1924b) is sympatric with Ecdyonurus in far western North America, but the larva is unknown and so diagnostic characters cannot be provided. In the eastern Palearctic, Nixe joernensis (Bengtsson, 1909) is sympatric with several species of the Ecdyonurus simplicioides group, but there is insufficient information to provide characters that will consistently differentiate Ecdyonurus and Nixe in this region.
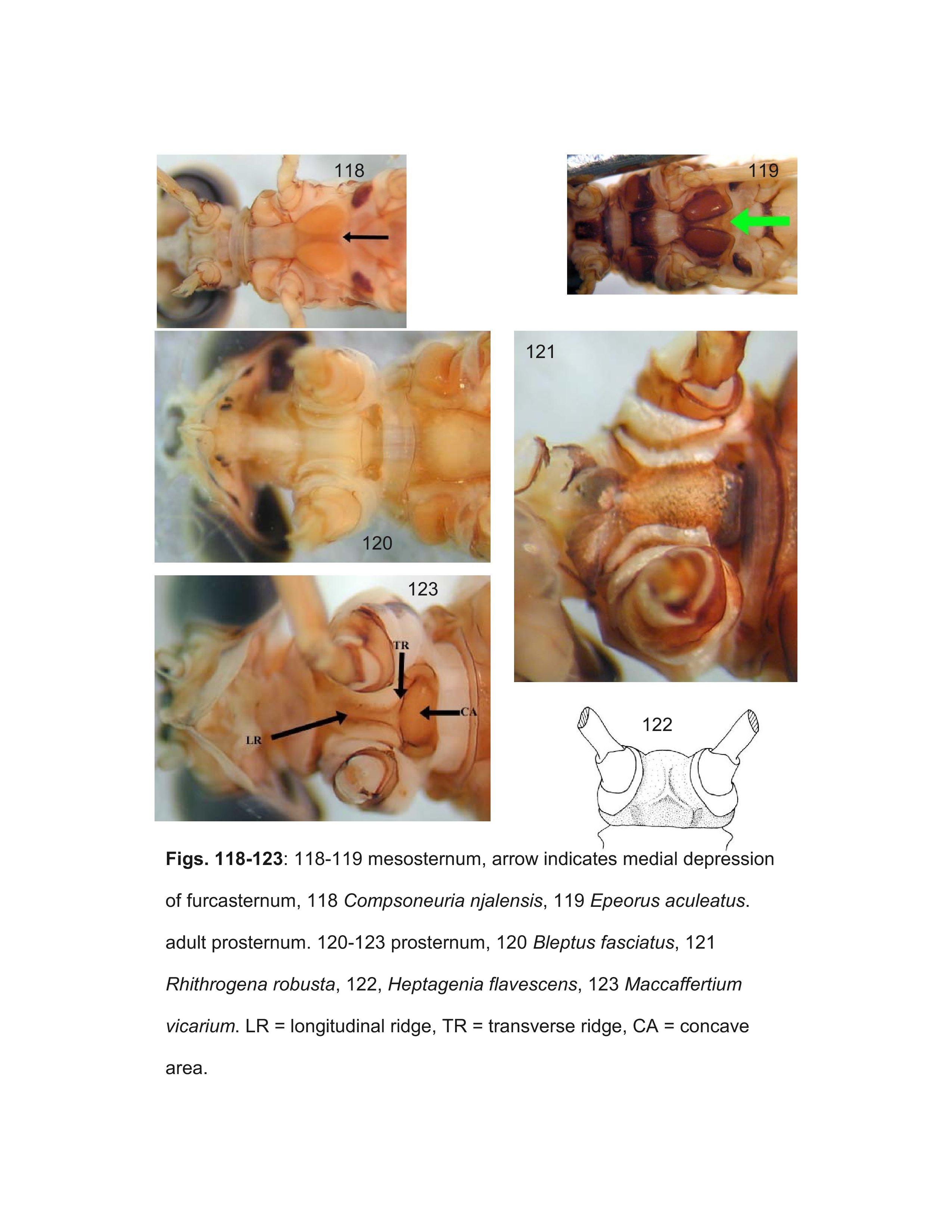
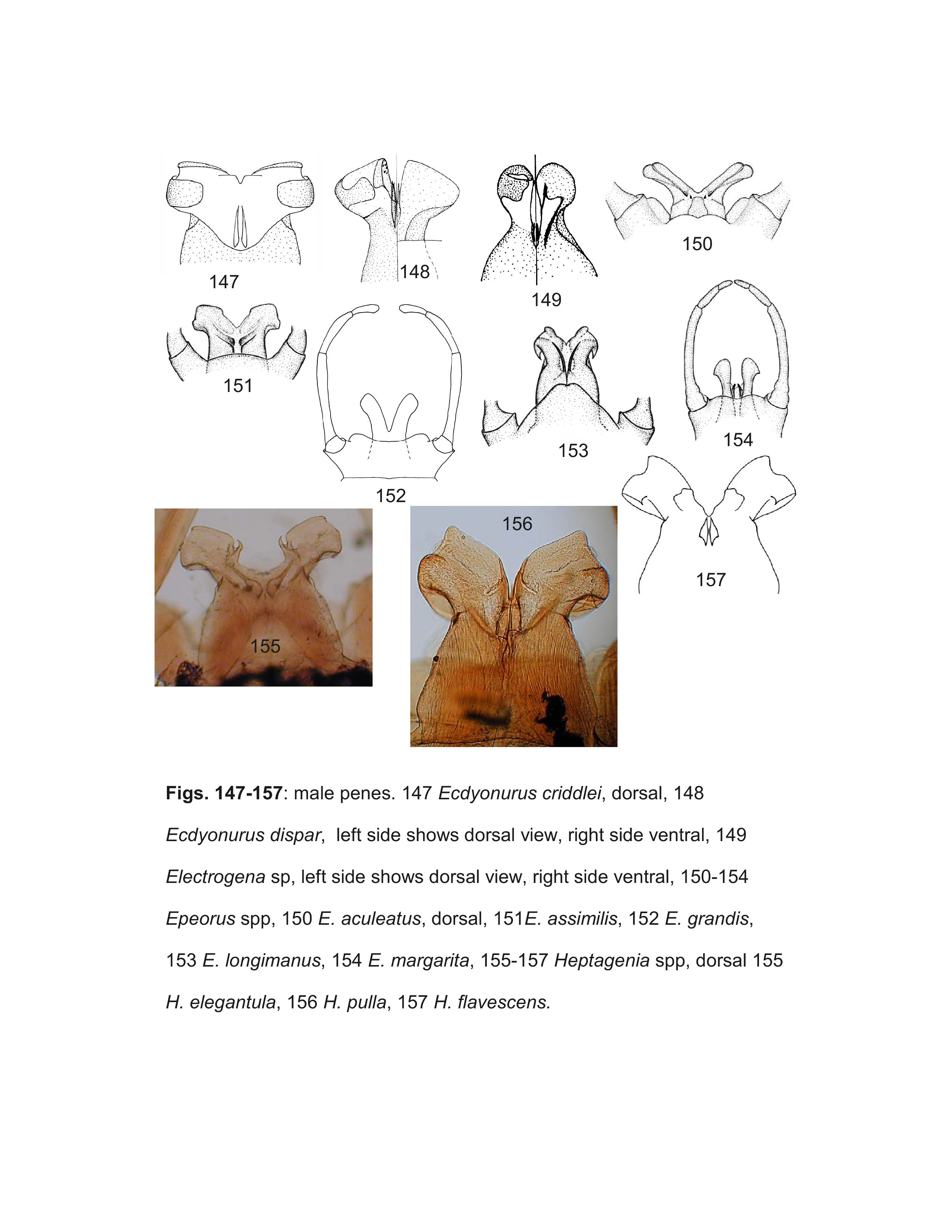
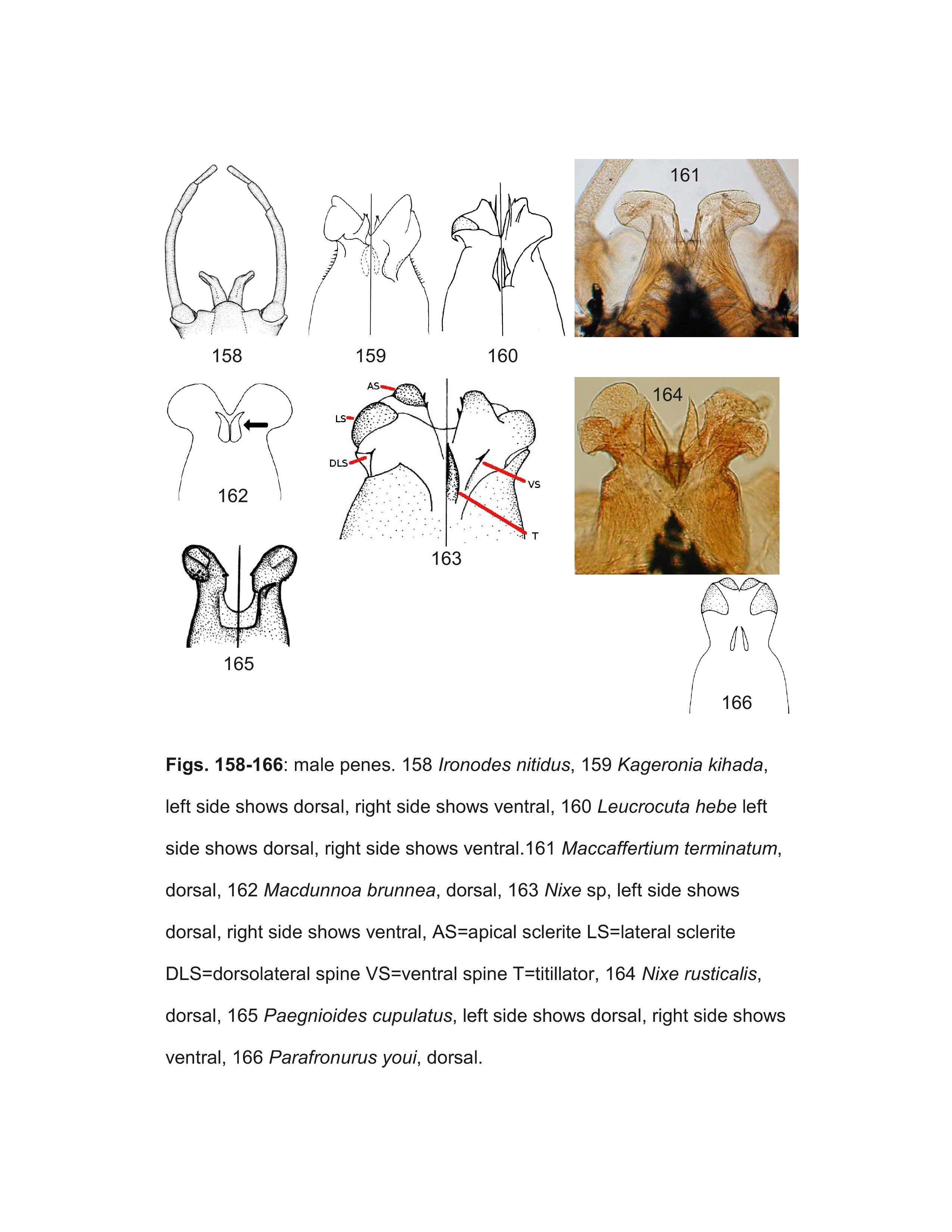
The key should correctly identify males of most genera, but some difficulties within the Ecdyonurinae may be encountered due to our lack of knowledge, particularly in the highly diverse and poorly known Oriental realm. The terminology used for the various spines, sclerites, and titillators of the penes are illustrated in Figure 163.
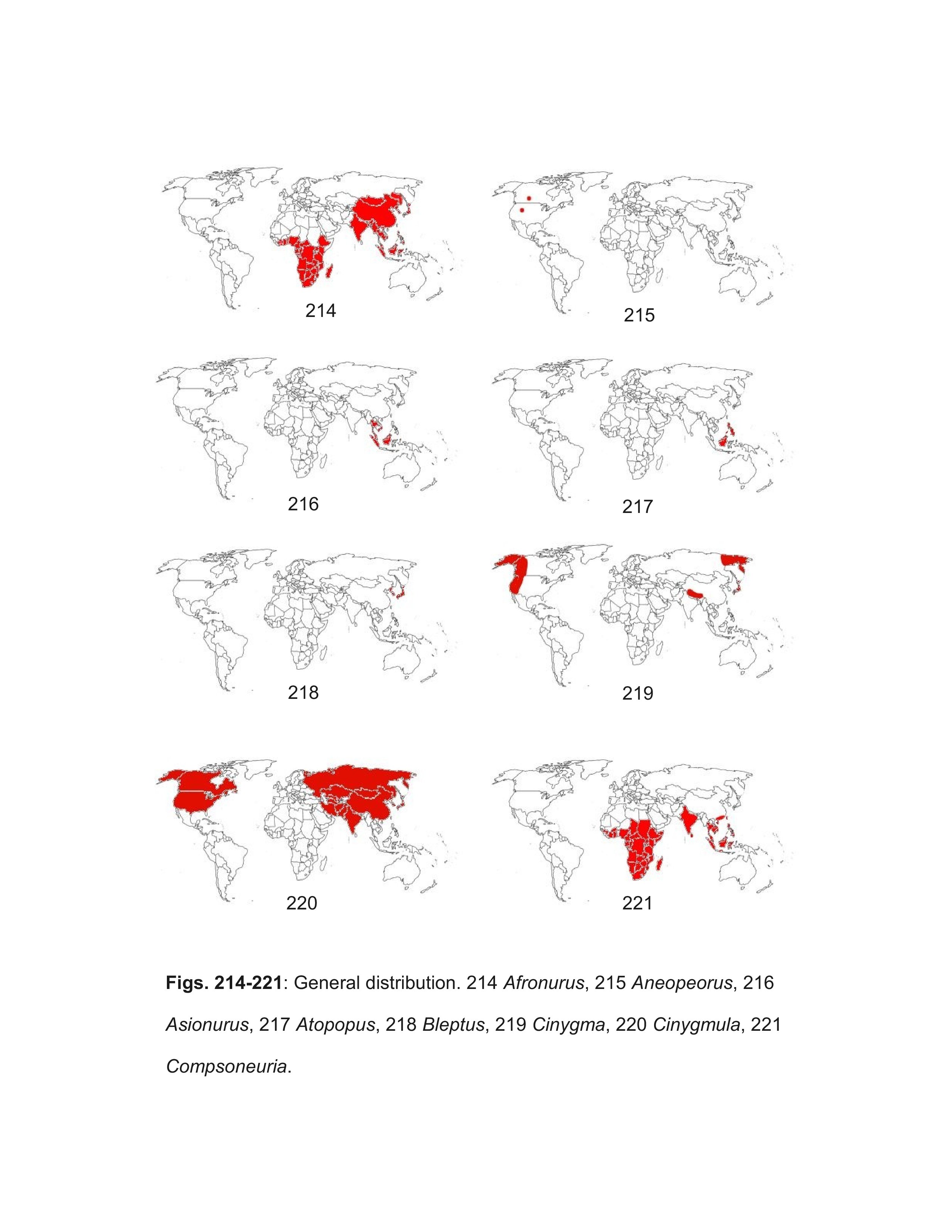
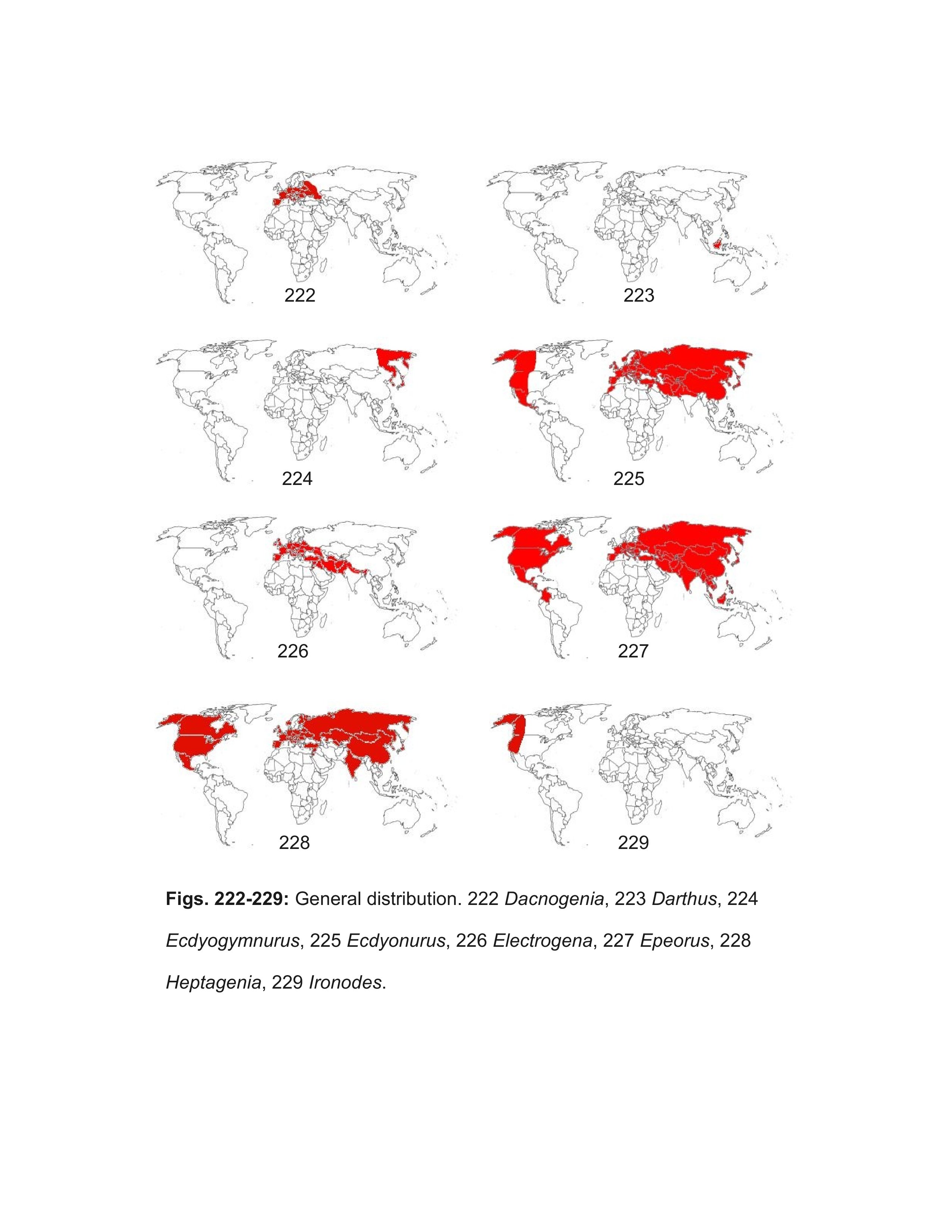
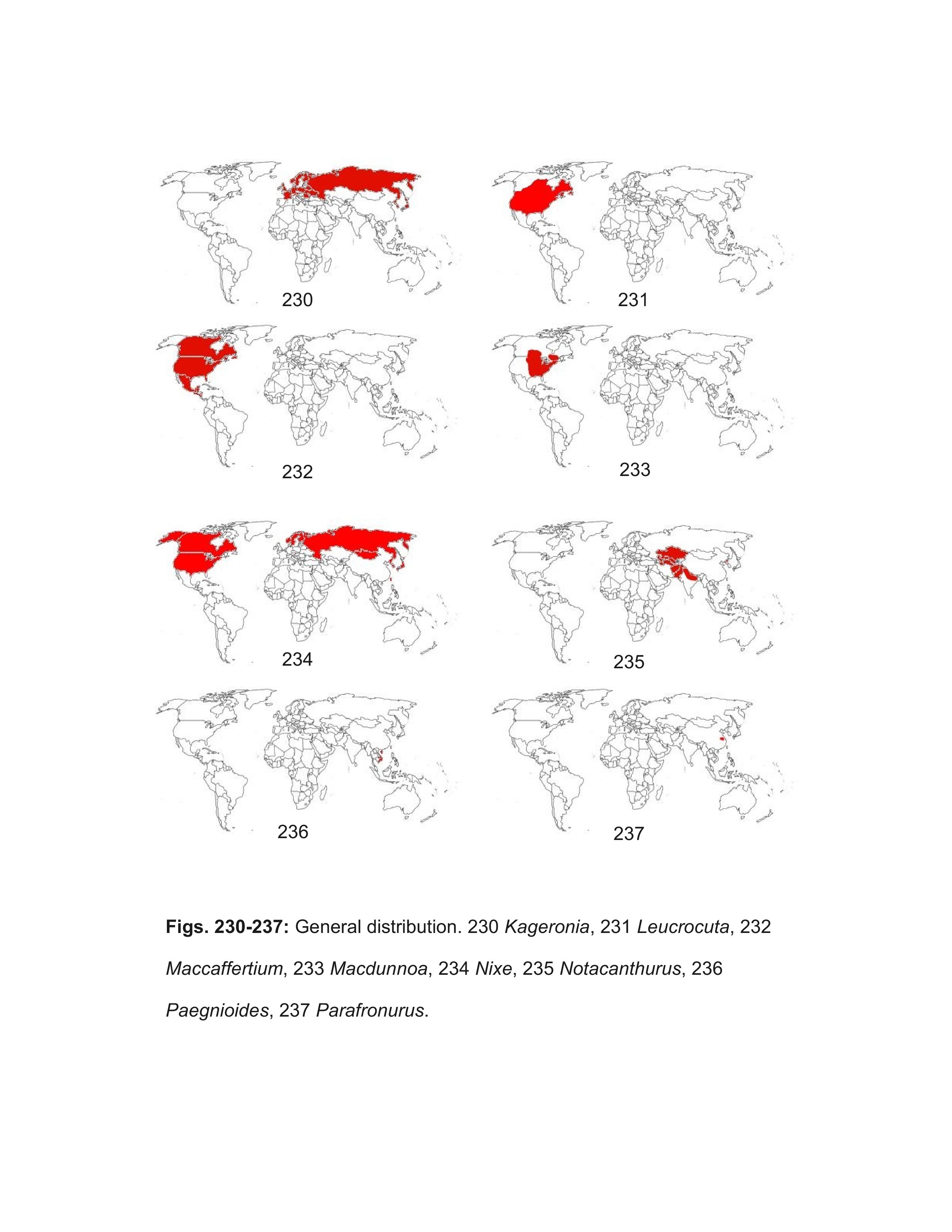
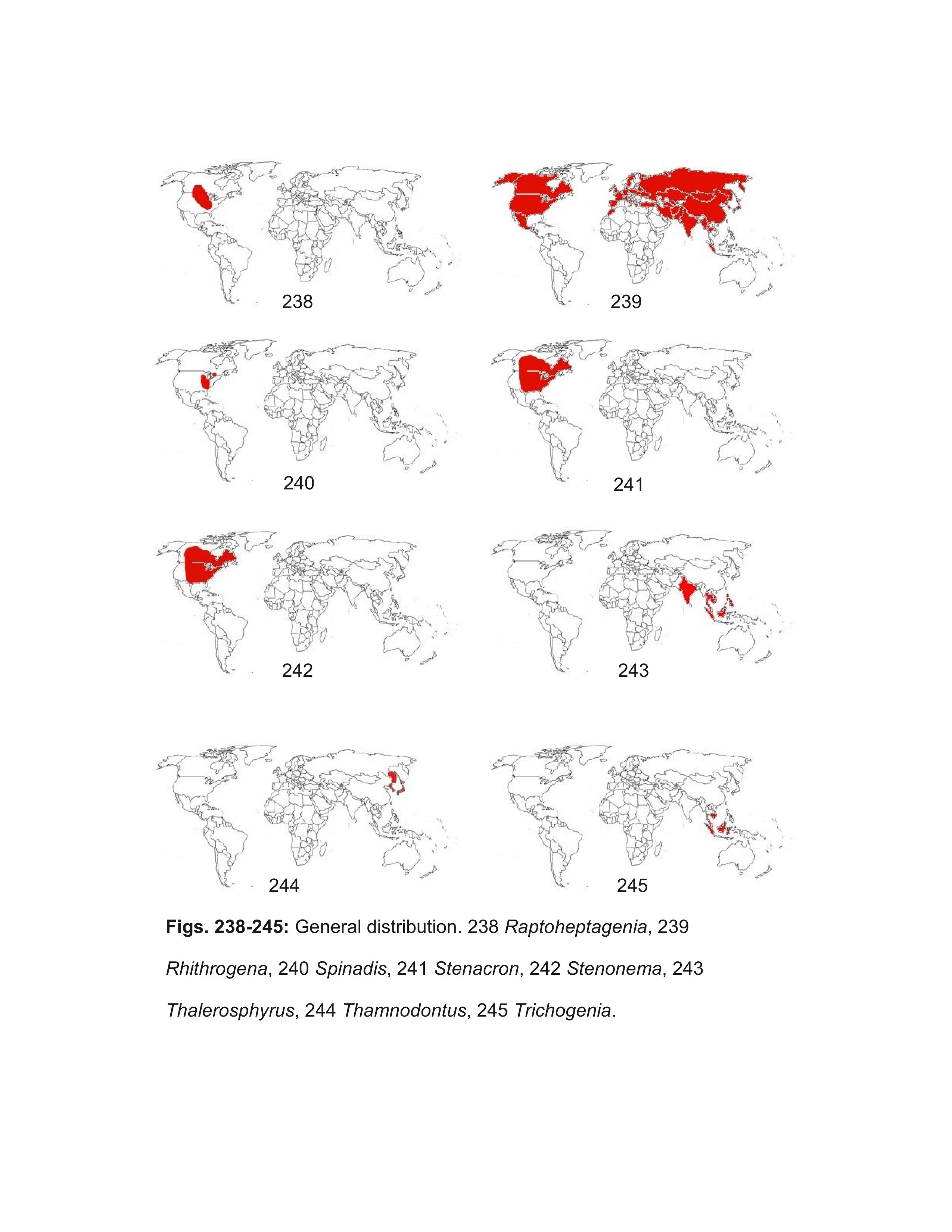
Most well marked North American females should be correctly identified using the key. Females of some of the Heptageniinae genera can be difficult to distinguish, but examination of eggs under magnifications of 400-1000X will confirm their identity. Distribution maps are referenced when their use will aid identification; general biogeographic zones are as in Figure 246. The following genera, arranged alphabetically under their respective subfamilies, are included in the male and larval keys:
- Ecdyonurinae
- Afronurus Lestage, 1924
- Asionurus Braasch & Soldán, 1986
- Atopopus Eaton, 1881
- Compsoneuria Eaton, 1881
- Darthus Webb & McCafferty, 2007 (Adult unknown)
- Ecdyogymnurus Kluge, 2004
- Ecdyonurus Eaton, 1868
- Electrogena Zurwerra & Tomka, 1985
- Leucrocuta Flowers, 1980
- Nixe Flowers, 1980
- Notacanthurus Tshernova, 1974
- Parafronurus Zhou & Braasch, 2003
- Thalerosphyrus Eaton, 1881
- Thamnodontus Kluge, 2004
- Heptageniinae
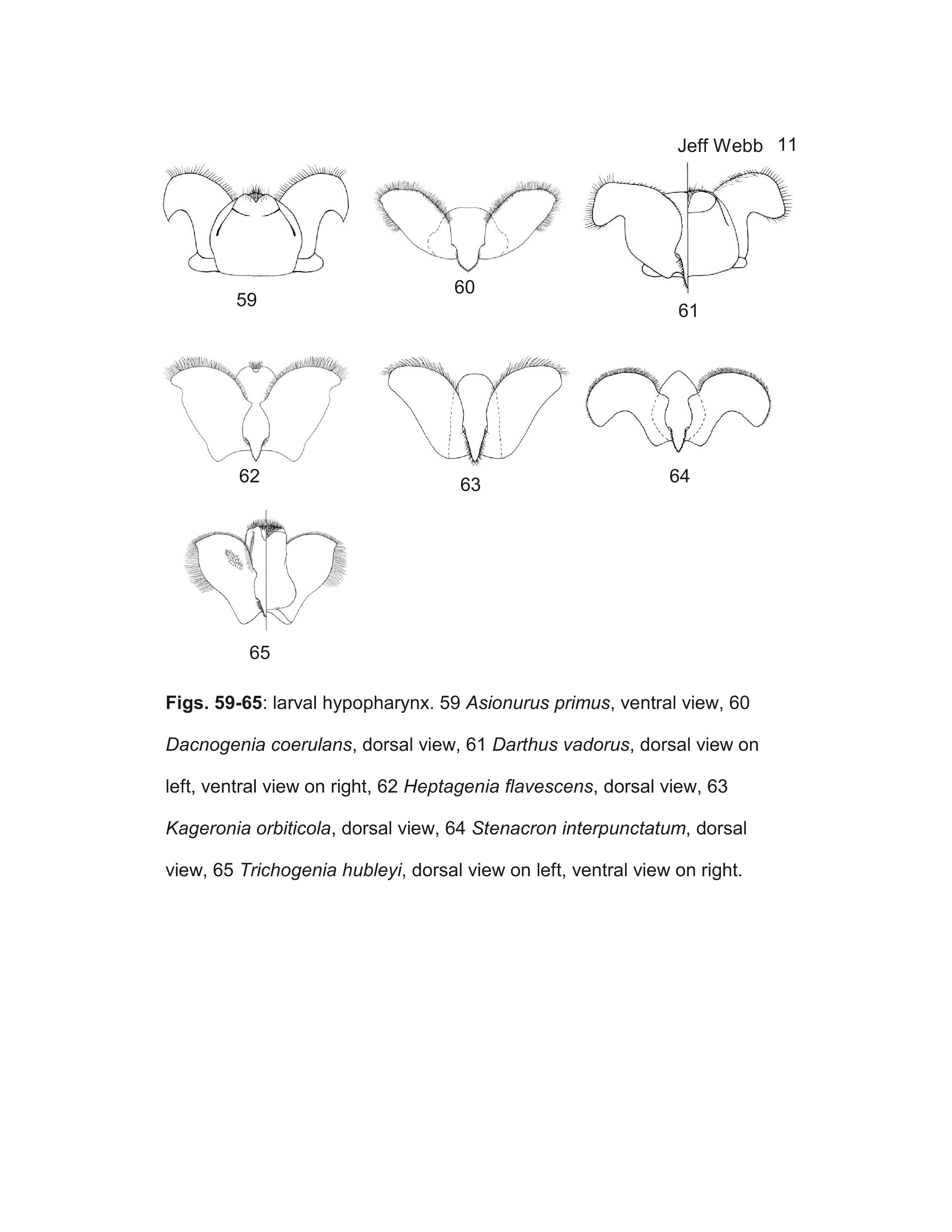
- Dacnogenia Kluge, 1988
- Heptagenia Walsh, 1863
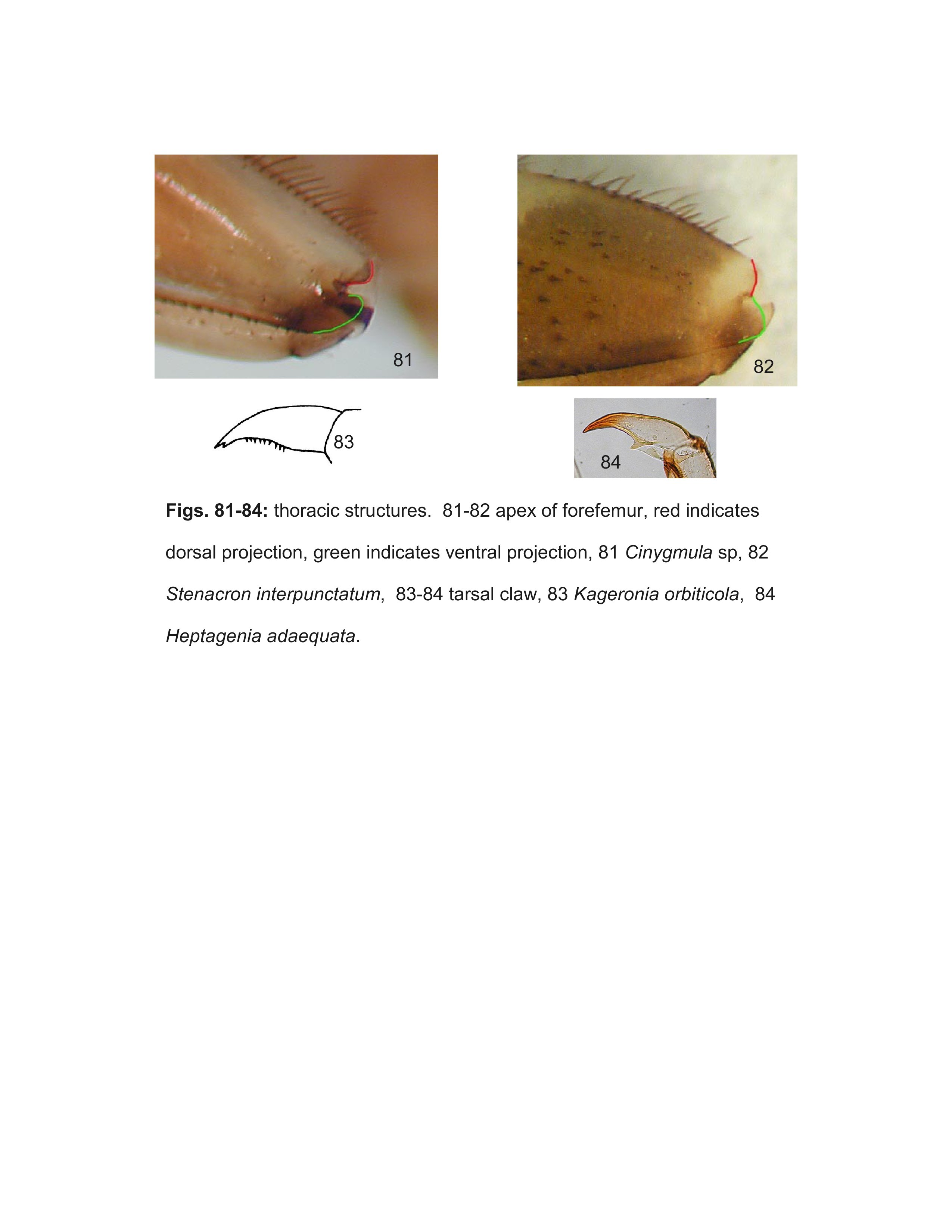
- Kageronia Matsumura, 1931
- Maccaffertium Bednarik, 1979
- Macdunnoa Lehmkuhl, 1979
- Raptoheptagenia Whiting & Lehmkuhl,1987
- Stenacron Jensen, 1974
- Stenonema Traver, 1933
- Trichogenia Braasch & Soldán, 1988
- Rhithrogeninae
- Anepeorus McDunnough, 1925
- Bleptus Eaton, 1885
- Cinygma Eaton, 1885
- Cinygmula McDunnough, 1933
- Epeorus Eaton, 1881
- Ironodes Traver, 1935
- Paegnioides Eaton, 1881
- Rhithrogena Eaton, 1881
- Spinadis Edmunds & Jensen, 1974

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}